
Michael Behe, Molecular Machines: Experimental Support for the Design Inference
Silmäsarja
Kuinka me näemme? Silmän anatomia tunnettiin jo 1800-luvulla hyvin yksityiskohtaisesti, ja monimutkaiset mekanismit, jotka tuottavat tarkan kuvan ympäröivästä maailmasta, ällistyttivät jokaista asiaan perehtynyttä ihmistä. 1800-luvun tiedemiehet havainnoivat, että jos ihmiseltä puuttui yksi silmän monista erikoispiirteistä kuten linssi, värikalvo tai silmälihakset, oli seurauksena vakava näön heikentyminen tai sokeus. Tästä tehtiin johtopäätös, että silmä voi toimia vain jos se on lähes vahingoittumaton.
Kun Charles Darwin mietti mahdollisia vastaväitteitä Lajien Synty-kirjassaan kohdistuen hänen teoriaansa evoluutiosta luonnonvalinnan avulla, hän käsitteli silmän ongelmaa kirjan osuvasti nimetyssä kappaleessa ”Organs of Extreme Perfection and Complication.” Hän ymmärsi, että jos yhdessä sukupolvessa ilmestyisi silmän monimutkaisuutta vastaava elin, kyseessä olisi suoranainen ihme. Jotta Darwinistinen evoluutio olisi uskottava, täytyi hänen jotenkin selvitä yleisön käsityksestä monimutkaisten elinten asteittaisesta kehityksestä.
Darwin keksi nerokkaan selityksen. Itse asiassa hän ei osoittanut miten evoluutio oli kehittänyt silmän, vaan hän osoitti vain, että on erilaisia eläimiä, joilla on rakenteeltaan erilaisia silmiä. Silmät vaihtelivat yksinkertaisista valonherkistä ”täplistä” monimutkaisiin selkärankaisten kamerasilmiin, ja näin ihmissilmän evoluutiossakin oli saattanut olla samantapaisia elimiä silmien välimuotoina.
Mutta kysymys on ennallaan, kuinka me näemme? Vaikka Darwin kykeni vakuuttamaan melkein koko maailman siitä, että moderni silmä oli voinut syntyä asteittain paljon yksinkertaisemmasta rakenteesta, hän ei edes yrittänyt selittää kuinka yksinkertainen valonherkkä ”täplä”, joka oli hänen lähtökohtansa, itse asiassa toimi. Silmästä puhuessaan Darwin sivuutti kysymyksen sen perimmäisestä mekanismista [1]:
Se kuinka hermo tulee valoherkäksi, tuskin kiinnostaa meitä enemmän kuin se, kuinka itse elämä sai alkunsa.
Hänellä oli erinomainen syy kieltäytyä vastaamasta kysymykseen: 1800-luvulla tiede ei ollut kehittynyt niin pitkälle, että kysymys olisi ollut edes käsiteltävissä. Kysymykseen: kuinka silmä toimii—se on, mitä tapahtuu, kun fotoni osuu verkkokalvoon—ei yksinkertaisesti pystytty vastaamaan tuohon aikaan. Itse asiassa, mihinkään kysymykseen, joka koski perimmäisiä elämän mekanismeja, ei pystytty vastaamaan. Kuinka lihakset saavat aikaan liikettä? Kuinka yhteyttäminen toimii? Kuinka energiaa saadaan ruuasta? Kuinka ruumis taistelee infektioita vastaan? Kukaan ei tiennyt.
Lassilaisuus
Kun tieto prosessin tarkasta mekanismista ei rajoita ihmismieltä, näyttää helpolta kuvitella yksinkertaisia askeleita jotka johtavat toimimattomasta toimivaan systeemiin. Hauska esimerkki tästä on suositussa sarjakuvassa Lassi ja Leevi. Pienellä Lassi-pojalla on useita seikkailuja tiikerinsä Leevin seurassa kun he hyppäävät laatikkoon ja matkaavat ajassa taaksepäin, nappaavat lelupyssyn ja ’transmuuntavat’ itsensä erilaisiksi eliöiksi, tai käyttävät laatikkoa kopioidessaan itseään selvitäkseen maailman voimista kuten äidistään ja opettajistaan. Pienelle lapselle kuten Lassi on helppoa kuvitella että laatikko todella voisi lentää kuten lentokone, koska Lassi ei tiedä miten lentokone toimii.
Hyvä esimerkki biologiasta maailmasta, jossa monimutkaiset muutokset näyttävät olevan yksinkertaisia, on usko spontaanista synnystä (spontaneous generation??). Yksi teorian puolustajista 1800-luvulla oli Ernst Haeckel, Darwinin suuri ihailija ja hänen teoriansa fanaattinen popularisoija. Sen rajallisen näkymän pohjalta, jonka 1800-luvun mikroskoopit tarjosivat soluista, Haeckel uskoi että solu oli ”yksinkertainen pieni kasa valkuaispitoista hiiltä,” [2] ei kovinkaan erilainen mikroskooppisesta hyytelöstä. Haeckelistä näytti siltä, että tuollainen yksinkertainen elämä voisi helposti syntyä elottomasta aineesta. Vuonna 1859, jolloin Lajien Synty julkaistiin, tutkimusalus H.M.S. Cyclops ruoppasi omalaatuista mutaa merenpohjasta. Nähdessään mudan Haeckel ajatteli sen muistuttavan paljon joitakin soluja jotka hän oli nähnyt mikroskoopissa. Innoissaan hän kertoi löydöstään Thomas Henry Huxleylle, joka oli Darwinin hyvä ystävä ja puolustaja. Myös Huxley vakuuttui että kyseessä oli Urschleimia, alkulimaa, itse elämän synnyttäjää, ja Huxley nimesi mudan Bathybius Haeckeliiksi huomattavan abiogeneesin puolustajan mukaan.
Muta ei kasvanut. Myöhemmin, biokemiallisten tekniikoiden ja mikroskooppien parantuessa, solun monimutkaisuus löydettiin. ”Yksinkertaisten kasojen” osoitettiin sisältävän tuhansia erityyppisiä orgaanisia molekyylejä, proteiineja, nukleiinihappoja, useita erillisiä rakenteita, erikoistuneiden prosessien erikoistuneita osastoja ja hyvin monimutkaisia rakenteita. Aikamme perspektiivistä katsoen Bathybius Haeckelii-episodi näyttää hassulta ja nololta. Mutta Haeckel ja Huxley käyttäytyivät luonnollisesti, kuten Lassi: koska he eivät tienneet solujen monimutkaisuudesta, heidän oli helppo uskoa että solu voisi syntyä yksinkertaisesta mudasta.
Läpi historian löytyy useita Haeckelin, Huxleyn ja solun kaltaisia esimerkkejä, jossa tieteellisen arvoituksen avainosa oli sen ajan ymmärryksen ulkopuolella. Tieteessä on jopa vitsikäs termi koneelle, rakenteelle tai prosessille joka tekee jotain, mutta tehtävän aikaansaavaa todellista mekanismia ei tunneta: sitä kutsutaan ’mustaksi laatikoksi’. Darwinin aikaan kaikki biologiassa oli mustaa laatikkoa: ei ainoastaan solu, silmä, ruuansulatus, immuniteetti, vaan kaikki biologiset rakenteet ja tuominnut koska, lopulta, kukaan ei kyennyt selittämään miten biologiset prosessit tapahtuivat.
Ernst Mayr, tunnettu biologi ja ohjaava voima uus-Darwinistisen synteesin takana, on todennut [3]:
Jokaisen tieteellisen vallankumouksen on hyväksyttävä kaikenlaisia mustia laatikkoja, sillä jos täytyisi odottaa kunnes ne kaikki on avattu, ei saavutettaisi mitään käsitteellistä edistystä.
Tämä on totta. Mutta aikaisempina aikoina kun mustia laatikkoja lopulta avattiin, tiede ja joskus koko maailma näyttivät muuttuvan. Biologia on edennyt merkittävästi Darwinin mallin seurauksena. Mutta Darwinin mustia laatikoita aletaan nyt avata ja kuvaamme maailmasta horjutetaan taas.
Proteiinit
Jotta voisimme ymmärtää elämän molekyyleihin nojaavan perustan, meidän täytyy ymmärtää miten proteiinit toimivat. Useimmat ihmiset pitävät proteiineja yhtenä ravintoryhmänä, eläimen tai kasvin kehossa proteiinit palvelevat eri tarkoitusta. Proteiinit muodostavat ovat elävän kudoksen koneiston, joka rakentaa tarpeelliset rakenteet ja saa aikaan elämälle tarpeelliset kemialliset reaktiot. Esimerkiksi proteiini Heksokinaasi saa aikaan ensimmäisen askeleen sokerin muuttamisessa biologisesti käyttökelpoiseen energiamuotoon. Iho muodostuu pitkälti kollageeni-proteiinista. Kun valo osuu silmäsi retinaan, se vuorovaikuttaa ensiksi rodopsin-proteiinin kanssa. Näemme näistä parista esimerkistä, että proteiinit suorittavat hyvin erilaisia toimintoja. Kuitenkin tietty proteiini pystyy yleensä vain yhteen tai pariin toimintoon: rodopsiini ei voi muodostaa ihoa eikä kollageeni reagoi hyödyllisesti valon kanssa. Siksi tyypillinen solu sisältää tuhansia ja tuhansia erityyppisiä proteiineja jotta se voi tehdä elämälle tarpeelliset tehtävät pitkälti samoin kuin työpajalla on useita työkaluja erilaisiin töihin.
Miltä nämä monipuoliset työkalut näyttävät? Proteiinien perusrakenne on melko samanlainen: Ne muodostetaan sitomalla yhteen aminohappoja. Vaikka proteiini rakenneketju voi pitää sisällään viidestäkymmenestä tuhanteen aminohappoa, jokainen paikka voi sisältää vain yhden kahdestakymmenestä aminohaposta. Näin proteiinit ovat kuten sanoja: sanat voivat olla eripituisia mutta ne koostuvat kirjaimista, joita on 26 erilaista. Proteiini ei loju solussa velttona ketjuna, vaan laskostuu hyvin tarkkaan muotoon, joka voi olla hyvin erilainen eri tyypin proteiineilla. Kaksi erilaista aminohappoketjua – kaksi erilaista proteiinia – voidaan laskostaa rakenteiksi, jotka ovat yhtä tarkoituksenmukaisia ja toisistaan erilaisia kuin kolmen tuuman jakoavain ja käsisaha. Ja kuten kodin työkalut, jos proteiinien muoto muuttuu merkittävästi, niiden toiminta lakkaa.
Ihmisen näkökyky
Molekyylitason biologiset prosessit tapahtuvat yleisesti ottaen proteiiniverkostojen avulla, joiden jokainen osa huolehtii tietystä tehtävästä.
Palatkaamme kysymykseen, miten näkeminen tapahtuu. Vaikkakin Darwinille näkemisen alkeistapahtuma oli täysi mysteeri, monien biokemistien työn tuloksena vastaus on tänään käden ulottuvilla. Kun valo saapuessa verkkokalvolle (retina), fotoni absorboituu siellä orgaaniseen molekyyliin nimeltä 11-cis-retinaali, joka tällöin muuntuu pikosekunneissa trans-retinaaliksi. Tämä muodonmuutos puolestaan pakottaa retinaaliin kiinteästi sitoutuneen rodopsiini-nimisen proteiinin vastaavaan 3-D-muodonmuutokseen. Proteiinin reaktiotapa muuttuu aivan erityisellä tavalla tämän metamorfoosin seurauksena. Muuntunut proteiini voi nyt reagoida erään toisen, transdusiini-nimisen proteiinin kanssa.
Ennen yhdistymistään rodopsiiniin transdusiini on tiukasti sidoksissa pieneen orgaaniseen molekyyliin, GDP:hen, mutta sitoutuessaan rodopsiiniin transdusiini irtoaa GDP:stä ja liittyy toiseen molekyyliin, GTP:hen, joka on GDP:lle sukua, mutta poikkeaa siitä oleellisesti.
GDP:n korvautuminen GTP:llä transdusiini-rodopsiini-kompleksissa muuttaa koko molekyylin käyttäytymistä. GTP-transdusiini-rodopsiini sitoutuu solukalvon sisäpinnalla olevaan fosfodiesteraasiin, joka puolestaan nyt sitouduttuaan kykenee kemiallisesti pilkkomaan molekyylin nimeltä syklinen GMP (cGMP). Prosessin alussa solussa on runsaasti cGMP:tä, mutta fosfodiesteraasin aktivoituminen alentaa sen pitoisuutta – kuten tapahtuu veden pinnalle kylpyammeessa, josta pohjatulppa vedetään irti.
Toinen cGMP:tä sitova solukalvoproteiini, ionikanava, voidaan kuvitella solun natriumionien määrää sääteleväksi käytäväksi. Se sallii normaalisti natriumionien pääsyn soluun, josta niitä pumppaa takaisin ulos erityinen proteiini. Solunsisäinen natriumpitoisuus pysyy hyvin tarkoissa rajoissa näiden kahden tekijän vuorovaikutuksen ansiosta. Kun nyt fosfodiesteraasi pilkkoo cGMP:tä ja alentaa sen pitoisuutta, ionikanavista useat sulkeutuvat, mistä seuraa solunsisäisten positiivisten natriumionien konsentraation lasku. Syntyvä jännite-ero solukalvon eri puolilla (siis hermosolun sähkövirta) etenee näköhermon aksonia pitkin aivoihin, joissa se tulkitaan näköaistimukseksi.
Jos tässä olisi koko näkötapahtuman biokemia, solusta loppuisivat nopeasti 11-cis-retinaali, cGMP ja myös natriumionit. Näköaistimuksesta vastaavien proteiinien lisäksi tarvitaan siis järjestelmä, joka rajoittaa aistimussignaalin kestoa ja palauttaa solun normaalitilaan. Tähän on solussa monia mekanismeja. Pimeässä ionikanava sallii natriumin lisäksi myös kalsiumionien pääsyn soluun; kalsium pumpataan sitten eri proteiinin toimesta takaisin oikean konsentraation ylläpitämiseksi. Kun cGMP-pitoisuus alenee sulkien ionikanavan ja alentaen Na-ionimäärää, myös Ca-ionimäärä alenee. Alentunut kalsiumpitoisuus puolestaan hidastaa merkittävästi fosfodiesteraasi-entsyymiä, joka tuhoaa cGMP:tä. Lisäksi guanylaattisyklaasi -niminen proteiini alkaa syntetisoida cGMP:tä uudelleen heti, kun kalsiumin pitoisuus lähtee laskuun. Tällä välin rodopsiinikinaasi -niminen entsyymi muuntaa metarodopsiini II:n kemiallisesti liittämällä siihen fosfaattiryhmän. Muuntunut rodopsiini sitoutuu arrestiini-proteiiniin, mikä sitoutuminen estää rodopsiinia aktivoimasta transdusiinia. Solulla on siis mekanismeja rajoittaa fotonin alullepanemaa vahvistettua signaalia.
Koska trans-retinaali irtoaa lopulta rodopsiinista, sen on seuraavaa valoaistimusta varten muunnuttava uudelleen 11-cis-retinaaliksi sekä jälleen sitouduttava opsiiniin muodostaakseen taas rodopsiinia. Kuitenkin erään entsyymin on ensin muokattava trans-retinaali kemiallisesti trans-retinoliksi liittämällä siihen kaksi uutta vetyatomia. Toinen entsyymi vielä isomerisoi molekyylin 11-cis-retinoliksi, ja lopuksi kolmas entsyymi poistaa aiemmin liitetyt kaksi vetyatomia, jolloin saadaan alkuperäinen molekyyli, 11-cis-retinaali, ja sykli on lopultakin valmis.
Elämän selittäminen
Vaikka kaikkia näkötapahtuman biokemian yksityiskohtia ei tässä ole mainittu, yleiskatsaus näkemisen fysiologiaan demonstroi hyvin sitä, mitä tarkoitetaan ”näkemisen selittämisellä”. Biologisen tieteen tulee lopulta selitykseen biokemiallisella tasolla. Jotta voitaisiin sanoa, että joku toiminto ymmärretään, jokainen olennainen askel prosessissa on selvitettävä. Biologiassa nuo askeleet ovat perustaltaan molekyylitasoisia, joten biologiaan kuuluvan ilmiön, kuten näkemisen, ruuansulatuksen tai immuniteetin, tyydyttävän selityksen osana on oltava molekyylitason selitys. Nyt kun näkötapahtuman musta laatikko on avattu, ilmiön riittäväksi”evolutiiviseksi selitykseksi” eivät enää riitä viittaukset kokonaisien silmien anatomiaan, kuten Darwin teki 1800-luvulla ja kuten useimmat evoluution popularisoijat tekevät edelleen. Anatomia on tässä, näkemisen perustapahtumassa, yksinkertaisesti epäolennainen. Samoin on fossiiliaineisto. Ei ole merkitystä, onko fossiiliaineisto yhdenmukainen evoluutioteorian kanssa, samoin kuin fysiikassa ei ollut merkitystä, olivatko Newtonin teoriat yhdenmukaisia arkipäivän kokemusten kanssa.
Fossiilit eivät kerro, miten 11-cis-retinaalin sekä rodopsiinin, transdusiinin ja fosfodiesteraasin väliset reaktiot ovat kehittyneet askel askeleelta. Myöskään biogeografisilla (eliöiden levinneisyyksien) malleilla ei ole tässä merkitystä, ei myöskään populaatiogenetiikalla tai niillä selityksillä, joita evoluutioteoria antaa surkastuneille elimille tai maapallon lajirunsaudelle.
”Kysymys, miten hermosolu alkaa aistia valoa, voisi tuskin koskea meitä enempää kuin kysymys siitä, miten elämä sai alkunsa”, sanoi Darwin 1800-luvulla. Kuitenkin moderni biokemia on kiinnostunut molemmista ilmiöistä. On mielenkiintoista havaita, miten elämän alkuperän tutkimus on hiljalleen halvaantunut, mutta tilanpuute estää kertomasta siitä tässä enempää. Riittää todeta, että tällä hetkellä elämän alkuperän tutkimus on hajaantunut ristiriitaisiksi selitysmalleiksi, joista jokainen on ei-vakuuttava, vakavasti puutteellinen ja yhteensopimaton kilpailevien mallien kanssa. Yksityisissä keskusteluissa jopa suurin osa evoluutiobiologeista myöntää että tieteellä ei ole selitystä elämän synnylle. [5]
Tämän artikkelin tarkoitus on näyttää, että samat ongelmat, jotka vaivaavat elämän alun tutkimusta, ovat myös esteenä, kun on yritetty osoittaa lähes minkä tahansa kompleksisen rakenteen syntymistä. Biokemia on paljastunut molekyylimaailman, joka itsepäisesti vastustaa kehitysteorian selitysyritystä, sen kehitysteorian, jota on kauan käytetty kokonaisten eliöiden tasolla. Kumpaakaan Darwinin mustista laatikoista – elämän alkuperää ja näkemisen perustapahtuman tai muidenkaan monimutkaisten biokemiallisen järjestelmän alkuperää – ei ole voitu selittää hänen teoriallaan.
Redusoitumaton monimutkaisuus
Lajien Synty-kirjassaan Darwin totesi [6]:

palautumattomasti monimutkaisesta järjestelmästä
“Jos voitaisiin näyttää toteen, että on olemassa jokin monimutkainen elin, joka ei mitenkään ole voinut muodostua lukuisien toisiaan seuraavien vähäisten muuntelujen tietä, olisi teoriani ehdottomasti kumottu.“
Järjestelmä, joka kohtaa Darwinin kriteerin on sellainen, joka on palautumattomasti monimutkainen. Palautumattomasti monimutkaisella tarkoitan yksittäistä järjestelmää, joka koostuu useista toisiinsa vaikuttavista osista, jotka osallistuvat perustoimintoon [perusfunktioon], ja jossa minkä tahansa osan poisto aiheuttaa käytännössä järjestelmän toiminnan lakkaamisen. Palautumattomasti monimutkaista systeemiä ei voi tuottaa graduaalisesti vähäisin, menestyksellisin muutoksin edeltävästä systeemistä, koska palautumattomasti monimutkaisen systeemin edeltäjä määritelmällisesti on ei-funktionaalinen. Koska luonnonvalinta edellyttää funktion valintaansa, palautumattomasti monimutkaisen systeemin – jos sellaista on – pitäisi olla ilmestynyt yhtenäisenä kappaleena, jotta luonnonvalinta voisi vaikuttaa siihen. Varsin yleisesti myönnetään, että sellainen yhtäkkinen tapahtuma olisi sovittamattomasti ristiriitainen Darwinin visioiman gradualismin kanssa. Tässä vaiheessa joka tapauksessa ”palautumattomasti monimutkainen” on vasta käsite, jonka teho on enimmäkseen sen määritelmässä. Meidän tulee kysyä, esiintyykö todellisuudessa palautumatonta monimutkaisuutta – ja jos esiintyy – niin, onko palautumattomasti monimutkaisia kohteita myös biologisissa järjestelmissä.
Pohditaan vaatimatonta hiirenloukkua (Kuva 1a). Hiirenloukut, joita perheeni käyttää kotonamme epätoivottujen jyrsijöiden pyydystämiseen, koostuu useista osista (Kuva 1b). Ne ovat: (1) Litteä alusta, joka toimii pohjana; (2) metallinen vasara, joka tekee varsinaisen työn pienen hiiren murskaamisessa; (3) Jousi levitetyin päin, joka pitää vasaran alustassa silloin, kun loukku on latingissa; (4) Herkkä liipasin joka laukaisee loukun kevyestä painosta; (5) Metallinen tanko (viritintanko), joka pitää vasaran takana kun loukku on viritetty ja yhdistetty liipasimeen. Erinäiset niitit ja ruuvit pitävät systeemiä koossa.

Jos jokin osa puuttuu, loukku ei toimi.
Jos jokin hiirenloukun komponenteista (alusta, vasara, jousi, liipasin, viritintanko) poistetaan, loukku ei toimi. Toisin sanoen, yksinkertaisella pienellä hiirenloukulla ei ole kykyä pyydystää hiirtä siihen asti, kuin vasta silloin kun useat erilliset osat on koottu yhteen.
Koska hiirenloukku on välttämättä koostuttava useista osista, se on palautumattomasti monimutkainen. Siten palautumattomasti monimutkaisia systeemejä on olemassa.
Molekyylikoneet
Onko redusoitumattoman monimutkaisia biokemiallisia systeemejä olemassa? Ehdottomasti. Monet entsyymikompleksit ovat sellaisia.
Aiemmin käsittelimme proteiineja. Monissa biologisissa rakenteissa proteiinit ovat suurempien molekyyli-koneiden alayksiköitä. Kuten kuvaputki, johdot, metallimutterit ja ruuvit muodostavat television, ovat monet proteiinit osa rakenteita jotka toimivat vain kun lähes kaikki komponenteista on asennettu. Hyvä esimerkki tästä ovat cilium-rakenteet.

Kuva 2a. Animaatio Ciliumista.
Ciliat ovat hiusta muistuttavia organelleja monien eläinten ja alempien kasvien solujen pinnalla. Ne siirtävät nesteitä solun pinnan yli tai ”soutavat” tahdissa yksittäisiä soluja nesteen läpi. Esimerkiksi ihmisissä hengitysteitä vuoraavilla epiteelisoluilla on aina noin 200 cilia-rakennetta per solu, jotka lyövät synkronisessa tahdissa, pyyhkien limaa kurkkua pitkin alas poistettavaksi. Cilium koostuu kalvon peittämästä kuitunipusta jota kutsutaan aksoneemiksi.Aksoneemi sisältää 9 tuplamikrotubuluksen renkaan joka ympäröi kahta keskellä sijaitsevaa yksittäistä mikrotubulusfilamenttia. Jokainen ulommaisista dubleteista koostuu 13 filamentin piiristä (alafilamentti A), jotka ovat fuusioituneet 10 filamentin yhdistelmäksi (alafilamentti B). Mikrotubulusten filamentit koostuvat kahdesta proteiinista, alfa ja beta tubuliinista. Aksoneemin muodostavat 11 mikrotubulusta pysyvät yhdessä kolmen tyyppisten konnektorien avulla: Alafilamentti A:t on yhdistetty keskisiin mikrotubuluksiin puolaproteiinilla säteittäin; vierekkäiset ulommista dubleteista on yhdistetty toisiinsa elastisen neksiini-proteiinin muodostamilla linkkereillä; lisäksi keskiset mikrotubulukset on yhdistetty sillan avulla toisiinsa. Viimein joka alafilamentti A kantaa kahta ”käsivartta”, sisempää ja ulompaa uloketta, joissa tavataan dyneiiniproteiini.
Mutta miten cilium-värekarva toimii? Kokeet ovat viitanneet siihen, että siliaarisen liikkeen salaisuus on kemiallisesta ”energiavaluutasta” voimansa saavassa tikku-ukon kävelyn kaltaisesta dyneiiniulokkeiden liikkeestä yhtä mikrotubulusta pitkin naapurissa sijaitsevaan alafilamentti B:hen siten että kaksi mikrotubulusta liukuvat toistensa sivuitse (kuva 2a ja b). Koskemattoman ciliumin mikrotubulusten väliset ristisidokset estävät kuitenkin vierekkäisiä mikrotubuluksia liukumasta toistensa sivuitse kuin pienen matkan. Nämä ristisidokset siten integroivat syneiinivetoisen liukuliikkeen kaareutuvaksi liikkeeksi koko aksoneemissa.
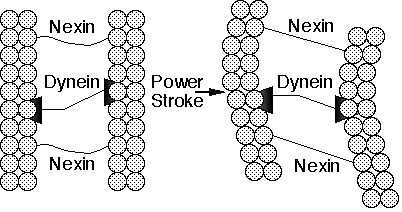
Kuva 2b. Kaaviopiirros ciliumista. Mikrotubuluksen moottoriproteiinin dyneiinin voima-isku naapuritubuluksen alafilamentti B:tä vasteen saattaa kuidut liukumaan toisiansa vasten. Joustava linkkeriproteiini neksiini muuttaa liukuvan liikkeen taipuisaksi liikkeeksi.
Luokaamme katsaus cilium-värekarvan toimintaan, pohtiaksemme mitä rakenne pitää sisällään. Ciliat ovat muodostuneet ainakin puolesta tusinasta proteiineja: alfatubuliinista, dyneiinistä, neksiinistä, puolaproteiinista, sekä keskisestä siltaproteiinista. Nämä yhtyvät funktioissaan toimittaakseen yhtä tehtävää, siliaarista liikettä, ja kaikkien näiden proteiinien täytyy olla läsnä jotta cilium toimisi. Jos tubuliinit ovat poissa, ei ole olemassa mitään filamenttia liukumaan. Jos dyneiinit ovat poissa, cilium pysyy jäykkänä ja liikkeettömänä. Jos taas neksiini tai jokin muu yhdistävä adapteri puuttuu, aksoneemi hajoaa teleskooppivavan lailla kun filamentit liukuvat.
Cilioissa emme siksi näe vain hämmästyttävää kompleksisuutta, vaan myös redusoitumatonta kompleksisuutta molekyylitasolla. Muistakaamme että ”redusoitumattomalla kompleksisuudella” tarkoitamme systeemiä joka vaatii useita erillisiä komponentteja jotta kokonaisuus toimisi.
Hiirenloukkuesimerkissäni täytyy olla pohja, vasara, jousi, loukku ja tukitanko, kaikki yhdessä toimien, jotta loukku toimittaisi tehtäväänsä. Aivan samoin ciliumissa, kun sellaista kasataan, täytyy olla liukuva filamentti, yhdistävät proteiinit ja moottorit jotta rakennelma toimittaisi tehtäväänsä. Jonkin komponentin puuttuessa koneisto on käyttökelvoton.
Ciliumin komponentit ovat yksittäisiä molekyylejä. Se tarkoittaa, ettei asiaan liity mitään ”mustia laatikoita”. Yksinkertaisimmankin värekarvan kompleksisuus on lopullista, ehdotonta. Ja aivan kuten tiedemiehet ja tutkijat, alkaessaan ymmärtää solun monimutkaisuutta, mielsivät miten hupsua olikaan ollut ajatella elämän nousseen spontaanisti yhdessä tai muutamissa askelissa valtamerten mudassa, niin mekin nyt ymmärrämme ettei yksinkertaisinta värekarvaammekaan ole voitu saada yhdessä tai muutamissa askeleissa itsestään. Koska värekarvan monimutkaisuus on redusoitumatonta, sillä ei voi olla toiminnallisia prekursoreita ja esimuotoja. Redusoitumaton kompleksisuus ei voi olla luonnonvalinnan tuote joka toimiakseen vaatisi funktioiden jatkumon josta valita. Valinta on voimaton kun ei ole mitään funktiota josta valita. Voimme mennä edemmäksi ja esittää väittämän, jonka mukaan värekarvan pitää olla suunniteltu jos sitä ei kerran voi tuottaa luonnonvalinnan avulla.
Molekyylien evoluution tutkimus
On monia muitakin esimerkkejä redusoitumattomasta monimutkaisuudesta. Näihin kuuluvat proteiinien siirtäminen, veren hyytyminen, suljettu rengasmainen DNA, elektroni siirto, bakteerin flagella, telomeronit, fotosynteesi, proteiinien koodaamisen säätely ja paljon muuta. Esimerkkejä redusoitumattomasta monimutkaisuudesta löytyy melkein joka sivulta biokemian oppikirjoista. Mutta jos näitä asioita ei voida selittää Darwinistisella evoluutiolla, miten tiedeyhteisö on ottanut tämän ilmiön vastaan viimeisen neljän vuosikymmenen aikana? Hyvä paikka etsiä vastausta tähän kysymykseen on Journal of Molecular Evolution -julkaisusarja. JME on lehti, joka perustettiin erityisesti käsittelemään molekyylitason evoluutiota. Sillä on korkeat tieteelliset vaatimukset ja sitä editoivat alan arvostetut nimet. Taannoisessa numerossa julkaistiin yksitoista artikkelia jotka kaikki käsittelivät proteiini- tai DNA sekvenssejä. Yksikään ei käsitellyt yksityiskohtaisia malleja välimuotoja monimutkaisten biokemiallisten rakenteiden kehittymiseksi. Viimeisen kymmenen vuoden aikana JME on julkaissut 886 artikkelia. Näistä 95 käsitteli proteiinien kemiallista synteesiä jonka ajatellaan olevan välttämättömiä elämän synnyn kannalta, 44 ehdotti matemaattisia malleja sekvenssi-analyysin parantamiseksi, 20 käsitteli nykyrakenteiden evoluutioteoreettisia seurauksia, ja 719 olivat proteiini ja polynukleotidi sekvenssejen analyysejä. Yksikään artikkeli ei käsitellyt yksityiskohtaisia malleja monimutkaisten biokemiallisten rakenteiden kehityksen välimuotoja. Tämä ei ole JME:n erikoisuus. Myöskään Proceedings of the National Academy of Science, Nature, Science ja Journal of Molecular Biology -lehdissä , eikä tietääkseni missään lehdellä, ei löydy artikkeleita jotka käsittelisivät yksityiskohtaisia kehityspolkuja.
Sekvenssien vertailut hallitsevat selvästi molekyylievoluution kirjallisuutta. Mutta sekvenssien vertailu ei yksinkertaisesti voi selittää monimutkaisten biokemiallisten systeemien kehitystä sen enempää kuin yksinkertaisten ja monimutkaisten silmien vertailu kertoi Darwinille miten ne toimivat. Tällä alueella tiede on mykkä. Tämä tarkoittaa sitä että kun päättelemme että monimutkaiset biologiset systeemit on suunniteltu, vastakkaista empiiristä näyttöä ei löydy, eikä ole vastakkaiseen viittaavaa teoreettista tutkimusta. Meidän ei tarvitse kyseenalaistaa yhtäkään empiiristä tulosta, mutta kokeiden tulkintaa täytyy tutkia tarkastaa samoin kuin Newtonin mekaniikan kanssa sopusoinnussa olevia kokeita piti tulkita uudelleen kun aalto-hiukkas-dualismi löydettiin.
Johtopäätökset
Usein sanotaan että tieteen täytyy välttää kaikkia johtopäätöksiä jotka viittaavat yliluonnolliseen. Minusta tämä on huonoa logiikkaa ja huonoa tiedettä. Tiede ei ole peli jossa käytetään mielivaltaisia sääntöjä päättämään mitkä selitykset hyväksytään. Pikemminkin se on yritys löytää tosia väittämiä todellisuudesta. Maailmankaikkeuden laajeneminen havaittiin vain kuusikymmentä vuotta sitten. Tämä tosiasia viittasi välittömästi erityiseen tapahtumaan – että jonain hetkenä menneisyydessä universumi alkoi laajeta hyvin pienestä koosta. Tämä päätelmä vihjasi vahvasti yliluonnolliseen tapahtumaan, luomiseen, universumin alkuun. Arvostettu fyysikko A.S. Eddington puhui todennäköisesti monien fyysikkojen puolesta puhuessaan tällaista vastaan [8]:
Filosifisesti nykyisen järjestyksen yht’äkkinen alku on minulle luotaan työntävä, kuten ajattelen se olevan monille, ja jopa ne jotka toivottaisivat Luojan asioihin puuttumisen todistuksen tervetulleeksi tulevat todennäköisesti ajattelemaan että yksi alkuunasettaminen joskus menneisyydessä ei juuri ole mieltä tyydyttävä suhde Jumalan ja hänen maailmansa välillä.
Big Bang kuitenkin hyväksyttiin fysiikkaan ja on vuosien kuluessa osoittautunut hedelmälliseksi paradigmaksi. Tärkeää on, että fysiikka seurasi dataa minne se näytti johtavan, vaikka jotkut ajattelivat että malli antoi apua ja lohtua uskonnolle. Nykyään kun biokemia löytää jatkuvasti esimerkkejä fantastisen monimutkaisista molekyylisysteemeistä, jotka lannistavat yritykset niiden synnyn selittämisestä, meidän tulisi ottaa oppia fysiikasta. Suunnittelun johtopäätös seuraa luonnollisesti mittausaineistosta, meidän ei tulisi väistää sitä, vaan hyväksyä se ja rakentaa sen päälle.
Lopuksi, on tärkeää ymmärtää, että emme päädy suunnitteluun sen perusteella, mitä emme tiedä, vaan sen perusteella, mitä tiedämme. Emme päädy suunnitteluun yrittäessämme selittää mustaa laatikkoa, vaan avointa laatikkoa. Primitiivisen kulttuurin edustaja saattaisi auton nähdessään arvata sen liikkuvan tuulen voimasta tai alle piilotetun antiloopin avulla, mutta kun hän avaa konepellin ja näkee moottorin, hän ymmärtää välittömästi että auto on suunniteltu. Samoin biokemia on avannut solun tutkiakseen mikä saa sen toimimaan ja me myös näemme että se on suunniteltu.
Viitteet
1. Darwin, Charles (1872) Origin of Species 6th ed (1988), p.151, New York University Press, New York.
2. Farley, John (1979) The Spontaneous Generation Controversy from Descartes to Oparin, 2nd ed, p.73, The Johns Hopkins University Press, Baltimore.
3. Mayr, Ernst (1991) One Long Argument, p. 146, Harvard University Press, Cambridge.
4. Devlin, Thomas M. (1992) Textbook of Biochemistry, pp.938954, WileyLiss, New York.
5. Washington yliopiston John Angus Campbell on huomauttanut ”Suuret idearakennelmat kuten positivismi eivät koskaan oikein kuole. Ajattelevat ihmiset hylkäävät ne asteittain ja jopa nauravat niille keskenään, mutta pitävät taivuttelussa käyttökelpoiset osat asiaan perehtymättömien käsittelyä varten.” The Comic Frame and the Rhetoric of Science: Epistemology and Ethics in Darwin’s Origin,” Rhetoric Society Quarterly 24, pp.2750 (1994). Tämä pätee selvästi siihen miten tieteellinen yhteisö käsittelee elämän synnyn kysymystä.
6. Darwin, p.154.
7. Voet, D. & Voet, J.G. (1990) Biochemistry, pp.11321139, John Wiley & Sons, New York.
8. Cited in Jaki, Stanley L. (1980) Cosmos and Creator, pp.56, Gateway Editions, Chicago.
Copyright ©1997 Michael Behe. All rights reserved. International copyright secured.